A dynamic CD8+ T cell-Tcell interaction may drive their differentiation into becoming protective immune cells and answer the age-old conundrum of CTL-based vaccine development and their inability to form central memory responses.
δ/αβ T cells and Ag recognition
Hi there.
My first blog entry and I've decided to start by talking about a recent paper I read and enjoyed, written by Pellicci et al. and published in JEM (2014). Here is a link to the original paper, The molecular bases of δ/αβ T-cell mediated Antigen recognition. I've always been a fan of Dale Godfrey papers, and the last few ones have definitely been top shelf. So, it makes sense to start with this paper.
Brief background:
Traditionally, αβ T cell receptors (TCRs) have been known to recognize antigens presented by Class I (in case of CD8) or Class II (CD4) MHC peptides that are expressed on antigen (Ag) presenting cells. In the past 10 years, evidence has been shown by many that suggests that certain T cell lineages are able to express lipid-based Ags from the CD1 family. Using Crystallography, Pellicci et al shows that the peculiar T cell subset expressing δ/αβ TCR is able to recognize CD1d-restricted-α-GalCer in a manner that is distinct from NKT cells. This is relevant because CD1d-α-GalCer is a glycolipid extracted and isolated from the marine sponge, and a prototypic ligand for activating type I NKT cells. For more information on NKT cells and CD1d, Godfrey papers are a great resource.
Central Hypothesis:
δ/αβ T cells are a group of cells "that bind Ag by way of both Vδ and Vβ genes, thus reflecting a greater level of diversity and functional potential withing T cell lineage."
Main questions addressed by the study:
- What is the frequency of δ/αβ T cells?
- How do they see an Antigen?
- What kind of Ag do they recognize?
- Can we coerce this interaction?
- How is the Ag recognition by δ/αβ TCR distinct from a conventional αβ TCR or a γδ TCR or Type I NKT cells?
The first part of the paper deals with characterizing the δ/αβ T cells using flow cytometry. Authors show that δ/αβ T cells are less frequent in the total pool from normal human PBMCs and highly variable from one donor to another. They have a hybrid TCR where a Vδ (δ1) chain is fused to Jα and a Cα parts of the α-chain, and pairs better with a β-chain (albeit, not expressing Vβ11 -characteristic of Type I NKT cells). This pairing is better than that of a γ-chain, which is expected for a γδ TCR. This is important, because the cytokine expression of IL-2, 4, 13, 17A, IFNγ, GM-CSF as well as TNF- is very similar to that of αβ T cells.
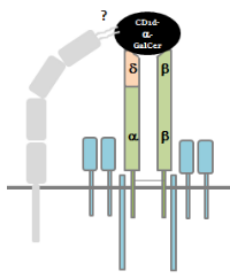
Second part of the paper deals extensively about the interaction of the δ/αβ TCR to CD1d-restricted-α-GalCer Antigen. Authors thoroughly examine the similarities and differences of this interaction by using specific clones for NKT cells, αβ T cells and γδ TCR. The intricacies of crystallography reveal that CD1d-α-GalCer interaction to the δ/αβ TCR is highly variable from clone to clone and within donors. However, in an example shown by the 9B4 clone (specific mAb for the δ/αβ TCR), authors clearly show that there is a 2-point interaction where both Vδ1 chain and the β-chain are involved, contributing to a better affinity and longer interaction time. To summarize, this small subset of T cells bearing a hybrid δ/αβ TCR is able to recognize a CD1d-α-GalCer antigen with high affinity.
Here are my reasons why this study is so fantastic and significant for the field:
For non-conventional T cells, such as those bearing a γδ TCR, there are very few known ligands and therefore, fewer ways of selective expansion. Having a system described by Pellici et al materializes ways in which γδ T cells can be selectively expanded and analyzed for requirements for their pairing γ-chains. This is especially important in studies which look at gene expression tied to specific TCR chains.
Beyond the expression of cytokines, Pellicci et al does not add much to the functional side of these T cells, which makes sense as the scope of the paper narrowly evaluates the requirements for ligand interaction. From the flow cytometry data, it seems that most of the δ/αβ Tcells in this study seem to coexpress DN or CD8+ T cells.
Generally, speaking, the most intriguing question is the function of δ/αβ T cells development of central memory and effector functions of T cells. Ag recognition and expansion is incomplete without understanding the subsequent transcriptional consequences. In other words, what other Ags can the dual δ and β - bound recognition system recognize?
Here's my most bold claim of all: is there a possible link to the most important riddle of T cell lineages.. a link between the lineage committment between the αβ and the γδ T cells? Yes, the α locus carries δ within and subsequently deletes δ upon formation of a fully functional α chain for the αβ TCR. Maybe these cells are generated and conserved with the hybrid receptor for a specific function, one which we do not know of ...not as of now!
Stay tuned for the next review that will pick up where we end.
Following is my graphical representation of the δ/αβ TCR based on the findings in this paper and an added modification of existence of an unknown co-receptor molecule expressed on these T cells.