Part-1 : The set up and the preliminary questions
Continuing the conversation about T cells with peculiar TCRs, I want to talk about CD4+ γδ Tcells. Specifically, how they are developed and how they change the committment and transform into αβ TCRs.
T cells never fail to intrigue me with their ability to differentiate and memorize the numerous antigenic signatures from a variety of pathogens and provide robust recall responses from second exposure (and onward). αβ T cells do a majority of the heavy lifting by compartmentalizing into functional phenotypic subsets such as naive, effecetor and memory T cells. Evaluation of plasticity or interchangeability of one functional phenotype of T cell to another is an ongoing area of research.
Hendrik Ziegler and colleagues recently published a scandalous paradigm shifting study in Frontiers of Immunology (Dec 2014) about a subset of T cells that co-expresses a CD4 co-receptor and a Vγ1+ γδ TCR. Here's a link to the Original study.
Relevant background
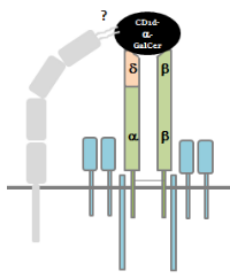
How T cells get those γ and δ chains or α and β chains, is a well characterized pathway. Below is a figure that highlights the stages of T cell development and the steps of maturation. There is a lot of detail that goes with the figure below, but this generalized version is much easier to look at.


To become a fully functional αβ TCR, genes for functionally rearranged β-chain are expressed leading upto the forking of T cell committment into CD4+ or CD8+. The thing to remember here is that the δ locus is embeded into the α locus and is deleted to form a fully functional αβ TCR to be formed.


In addition to the depletion of functional thymus through our lifetime (as shown in the bargraph above), "The thymic involution begins as early as 1 year after birth, resulting in an exponentially decreasing output of naïve T cells, which is almost completely extinguished post adolescence. The total size of the T-cell pool nevertheless remains relatively constant throughout life, which suggests that the T-cell pool must be replenished in some other way." (From Ziegler et al.) How does this happen has been studied extensively through past few decades, which instigates whether the plasticity among the different lineages such as γδ vs. αβ may also contribute in replenishment of the naïve T cells. What is needed, however, is a pool of cells with stem cell markers that can differentiate into lineages starting from the pre-TCR phenotype.
Ziegler et al shows a mechanism for CD4 expressing γδ TCRs that carry markers for earliest hematopoietic progenitor cells and carry full-length transcripts of in-frame δ, γ, and β TCR gene rearrangements in addition to genes and enzymes that are characteristic of progenitor populations (such as DN1 and DN2). The central hypothesis of this study is as follows:
Under inflammatory environment, CD4+ γδ T cells can form fully functional αβ TCR+ T cells.
How this happens will be discussed in the subsequent parts of this blog.