
A dynamic CD8+ T cell-Tcell interaction may drive their differentiation into becoming protective immune cells and answer the age-old conundrum of CTL-based vaccine development and their inability to form central memory responses.
γδ TCR--> αβTCR: (Part3)
Part-3: Trans-differentiation of the TCR of CD4+ Vδ1+ γδT-Cell Clones into αβTCR
This part of the series will aim to discuss the detailed developmental steps of the trans-differentiation of CD4+ Vδ1+ γδT-cells to fully functional αβ T cells.
The CD4+ Vδ1+ γδT-cells clones were reportedly extremely long-lived and had a downregulated expression of CD34. In the clones that continued to grow with Vδ1+ γδTCRs, authors tracked the commitment change to αβTCR over one month. At day 7, the initially Vδ1+ γδT-cells that were negative for αβTCR, now started to express an αβTCR and became low on γδTCR. This expression profile changed entirely on day 28, where the cells became αβTCR(+) γδTCR(lo). The CD4 and CD8 expression on these clones was also evaluated and it turned out that the cells remained either SP CD4+ or changed their co-receptor from SP CD4+ --> DP CD4+/CD8dim. This resulted into cell pool containig either SP CD4+ or SP CD8+ αβTCR(+) T cell clones. Original figure (5) from the paper shows more details on staining.
Developmental steps leading to trans-differentiation
Expression of the constant region of TCR chains was used as a focal point whilst evaluating the re-organization of these clones. The chronological steps in which this change occurs is as follows:
- CD34 is down-regulated
- DN1-stage like Vδ1+ Cγ1+ Vδ1+ γδT-cells express CD4 and undergo morphological changes during proliferation
- Cells now become DN2-stage like where they are still αβ negative, but lower in expression of both Vδ1+ and Cγ chains, and start expressing Cβ chain. At this point cells have an LGL like proliferation profile
- Expression of the β-chain leads to progression towards DN3-stage and thereby further downregulation of the Vδ1 chain and ablation of Cγ chain replaced by a Cβ chain altogether
- Cells undergo β-chain selection with a fully formed Cβ chain, cells undergo DP-like developmental stage, where the expression of CD4+CD8+ αβTCR is dim but the competitive binding of Vβ1+Vδ1 vs. Vβ+Vα+ begins.
- At this point, the processive α-chain rearrangement occurs that deletes the δ-chain and leads to positive selection and form a DPb stage and express Ca+Cb+Vd1-Cg-
- Finally, the SP TCRαβ+ cells yield CD4+ or CD8+ αβ+TCR cells.
This summary can be shown graphically in the following summary figure extracted from the paper.

I adore this paper. It is not an easy paper to read with many critical and meticulous experimental details, but if you were to gloss over them and just focus on the things I did, you might find:
If, under severe inflammatory conditions, a mature Vδ1+ γδ TCR can undergo gene re-organization and form a fully functional CD4+ or CD8+ αβ TCR, that not only changes our convention of T-cell lineage but also build on the existsing perspective of the plasticity of these re-arrangements.
As a γδ T cell scientist, I always enjoy a paper that shows ways to expand and functionally characterize the non-conventional T cells in humans. While, some schools of thoughts still categorize γδ T cells as innate like lymphoid cells (ILCs), there are many that consider γδ T cells to be a member of the adaptive arm. Me, on the other hand, would go even a step further and put them in yet another category of "bridging immunity". While, γδ T cells are first to respond at sites of inflammation in many models and are able to expand in recall responses, Ziegler shows that they are also able to trans-differentiate into fully functional αβTCRs.
Further experiments need to be done in different model systems to truly assure that the clonal reorganization of TCR commitments is changing and not just an artifact. Ziegler et al does a fantastic job in their discussion by validating the findings by comparing them to evidences put forth by other papers in the last couple decades.
It would also be interesting to see this sort of a model for gene re-organization in more severe infectious models such as HIV and malignant tumors. One wonders if the tumor microenviroment poses such reorganization and if so, does that make the tumor malignancy more severe?
Please feel free to clarify points that I may have missed or misinterpreted.
γδ TCR--> αβTCR (Part 2)
Built on the foundation of last entry's T cell lineage commitment summary, this part will focus on the specifics of observed characteristics of heterodimeric TCR bearing (γδ/αβ) clones of CD4+ Vδ1+ γδ T cells.
Part-2: Characteristics of CD4+ Vδ1+ γδ T cell clones
Ziegler and colleagues were able to make extremely long-lived clones of CD4+ Vδ1+ γδ T cell clones by culturing lymphocytes from healthy individuals and observed the following:
- Clones were able to change their TCRs from γδ to αβ over time
- Clone morphology was similar to that of Large granular lymphocytes (LGLs)
- Constant region of the TCR-γ9+ chain contained a Cγ1 region
- Clones expressed CD34(lo), CXCR4, TGF-β and receptor, IL-7R, c-Kit and FLT-3.
- Like DN1-stage, CD4+ Vδ1+ γδ T cell clones were CD34+ (albeit low) CD38+ CD1a-.
- Clones rapidly expressed CD2 during cultivation, which was initially absent
- Spontaneously produced cytokines such as TGF-β, IL-2, -4, -5, -6, -10, -13, -17A, IFN-γ and TNF-α upon PMA+IONO
- Clones were CD45RO+, CD45RA-, CDG2L-, CD27- and CCR7-.. and therefore, characterized as effector memory cells.
Let's look at a couple of these in more detail.
(3) is interesting where they found that the C-region of TCR-γ9+ chain employs a Cγ1 region.
There are two major subpopulations of γδ T cells based on their "V" and "C" gene segment usage. In the PBMCs, majority of γδ TCRs are Vγ9/Vδ2. TCR of these chains link the γ and the δ chains via di-sulfide linkages. Therefore, the TCRγ and chains use the Cγ1 region and not Cγ2. Salvatore Ciccarese and colleagues wrote this beautiful paper comparing the differences in Cγ regions by the γδTCRs in three species: humans, mice and artiodactyls. Study is titled, "Evolution of T-cell Receptor Gamma and Delta Constant Region and Other T-Cell-Related Proteins in the Human-Rodent-Artiodactyl Triplet." In humans, he says, the main differences between the Cγ1 and Cγ2 is simple: there are 3 exons for Cγ1 and four for Cγ2. The second exon is duplicated in the Cγ2 which lacks a Cysteine that makes a di-sulfide bridge, and hence, Cγ1 is a major segment used by the TCRγ9 chains.
(8) Clones were likely effector memory cells. This is by far the most interesting characteristic because these clones are extremely long lived and are isolated from healthy human PBMCs. Ziegler et al states that these are not naiive but are effector memory cells. The reason this is the case is because they did not express the CD45RA Ag, suggesting they are not naive. To clarify, CD45RA is expressed on naïve T cells, as well as the effector cells in CD4+ populations. However, upon Ag experience, central (Tcm) and effector (Tem) memory T cells gain expression of CD45RO and lose expression of CD45RA. Therefore, we use either CD45RA or CD45RO to generally differentiate the naïve from memory populations. Another way to further differentiate naiive from effector or central memory is by quantitating the expression of CCR7 or CD62L. While both naiive and central memory cells expresss CCR7 and CD62L, effector memory or effector cells tend not to.
CD4+ Vδ1+ γδ T cell clones expressed CD45RO+, but not CD45RA, CD62L, CD27, or CCR7− and sometimes expressed CD28. While they lacked the CCR7 expression, the clones expressed other chemokine receptors such as CCR4, CXCR1/CXCR2, CCR6, and CXCR4 - which direct the movement of circulating T cells to sites of inflammation and tissue injury. This is important detail to remember and ties back the inflammatory microenvironment of the ex vivo cultures. Ziegler et al shows that the CD4+ Vδ1+ γδT cells
that lack thymus-homing properties but carry chemokine receptors (CCR) that direct circulating T cells to sites of inflammation, can develop into functional, mature CD4+ or CD8+ αβT cells in an inflammatory environment.
Individual steps of this development will be discussed in part 3.
δ/αβ T cells and Ag recognition
Hi there.
My first blog entry and I've decided to start by talking about a recent paper I read and enjoyed, written by Pellicci et al. and published in JEM (2014). Here is a link to the original paper, The molecular bases of δ/αβ T-cell mediated Antigen recognition. I've always been a fan of Dale Godfrey papers, and the last few ones have definitely been top shelf. So, it makes sense to start with this paper.
Brief background:
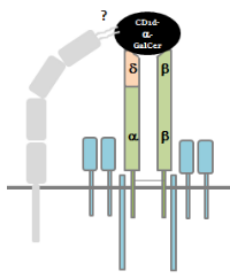
Traditionally, αβ T cell receptors (TCRs) have been known to recognize antigens presented by Class I (in case of CD8) or Class II (CD4) MHC peptides that are expressed on antigen (Ag) presenting cells. In the past 10 years, evidence has been shown by many that suggests that certain T cell lineages are able to express lipid-based Ags from the CD1 family. Using Crystallography, Pellicci et al shows that the peculiar T cell subset expressing δ/αβ TCR is able to recognize CD1d-restricted-α-GalCer in a manner that is distinct from NKT cells. This is relevant because CD1d-α-GalCer is a glycolipid extracted and isolated from the marine sponge, and a prototypic ligand for activating type I NKT cells. For more information on NKT cells and CD1d, Godfrey papers are a great resource.
Central Hypothesis:
δ/αβ T cells are a group of cells "that bind Ag by way of both Vδ and Vβ genes, thus reflecting a greater level of diversity and functional potential withing T cell lineage."
Main questions addressed by the study:
- What is the frequency of δ/αβ T cells?
- How do they see an Antigen?
- What kind of Ag do they recognize?
- Can we coerce this interaction?
- How is the Ag recognition by δ/αβ TCR distinct from a conventional αβ TCR or a γδ TCR or Type I NKT cells?
The first part of the paper deals with characterizing the δ/αβ T cells using flow cytometry. Authors show that δ/αβ T cells are less frequent in the total pool from normal human PBMCs and highly variable from one donor to another. They have a hybrid TCR where a Vδ (δ1) chain is fused to Jα and a Cα parts of the α-chain, and pairs better with a β-chain (albeit, not expressing Vβ11 -characteristic of Type I NKT cells). This pairing is better than that of a γ-chain, which is expected for a γδ TCR. This is important, because the cytokine expression of IL-2, 4, 13, 17A, IFNγ, GM-CSF as well as TNF- is very similar to that of αβ T cells.
Second part of the paper deals extensively about the interaction of the δ/αβ TCR to CD1d-restricted-α-GalCer Antigen. Authors thoroughly examine the similarities and differences of this interaction by using specific clones for NKT cells, αβ T cells and γδ TCR. The intricacies of crystallography reveal that CD1d-α-GalCer interaction to the δ/αβ TCR is highly variable from clone to clone and within donors. However, in an example shown by the 9B4 clone (specific mAb for the δ/αβ TCR), authors clearly show that there is a 2-point interaction where both Vδ1 chain and the β-chain are involved, contributing to a better affinity and longer interaction time. To summarize, this small subset of T cells bearing a hybrid δ/αβ TCR is able to recognize a CD1d-α-GalCer antigen with high affinity.
Here are my reasons why this study is so fantastic and significant for the field:
For non-conventional T cells, such as those bearing a γδ TCR, there are very few known ligands and therefore, fewer ways of selective expansion. Having a system described by Pellici et al materializes ways in which γδ T cells can be selectively expanded and analyzed for requirements for their pairing γ-chains. This is especially important in studies which look at gene expression tied to specific TCR chains.
Beyond the expression of cytokines, Pellicci et al does not add much to the functional side of these T cells, which makes sense as the scope of the paper narrowly evaluates the requirements for ligand interaction. From the flow cytometry data, it seems that most of the δ/αβ Tcells in this study seem to coexpress DN or CD8+ T cells.
Generally, speaking, the most intriguing question is the function of δ/αβ T cells development of central memory and effector functions of T cells. Ag recognition and expansion is incomplete without understanding the subsequent transcriptional consequences. In other words, what other Ags can the dual δ and β - bound recognition system recognize?
Here's my most bold claim of all: is there a possible link to the most important riddle of T cell lineages.. a link between the lineage committment between the αβ and the γδ T cells? Yes, the α locus carries δ within and subsequently deletes δ upon formation of a fully functional α chain for the αβ TCR. Maybe these cells are generated and conserved with the hybrid receptor for a specific function, one which we do not know of ...not as of now!
Stay tuned for the next review that will pick up where we end.
Following is my graphical representation of the δ/αβ TCR based on the findings in this paper and an added modification of existence of an unknown co-receptor molecule expressed on these T cells.
{kind=link}